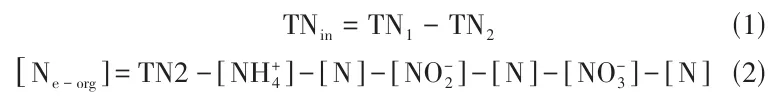一株寡养单胞菌属菌L12在高La(Ⅲ)浓度下的SND降解特性
1太原理工大学环境与生态学院,山西 太原 030024;2太原理工大学化学与化工学院,省部共建煤基能源清洁高效利用国家重点实验室,山西 太原 030024
陈虎, 翟春鹏, 王莹, 等. 一株寡养单胞菌属菌L12在高La(Ⅲ)浓度下的SND降解特性[J]. 化工进展, 2026, 45(4): 2409-2420.
>>DOI:10.16085/j.issn.1000-6613.2025-0583
生物脱氮是目前应用最广泛的氮污染控制方法,然而稀土矿山废水中除含有大量的含氮物质外,还含有大量的稀土元素离子。为此,本文从废水中分离筛选出能够耐La(Ⅲ)的同步硝化反硝化菌L12,通过菌落形态学和16S rDNA基因序列同源性分析,鉴定其归属于寡养单胞菌属(Stenotrophomonas sp.)。然后通过单因素实验优化了菌株L12的氨氮去除性能,结果表明菌株L12在含La(Ⅲ)(浓度为10mg/L)环境中利用氨氮的最佳条件为丁二酸钠为碳源、C/N=12、30℃和pH=8.0,可以在50~500mg/L的氨氮负荷下生长。菌株L12在初始La(Ⅲ)浓度为0~25mg/L环境下可以迅速生长和转化氨氮。但当La(Ⅲ)浓度大于25mg/L时,菌株L12生长缓慢,对氨氮的去除时间明显延长。菌株L12能够分别以氨氮、硝态氮和亚硝态氮为唯一氮源进行生长,对应的氨氮、硝态氮和亚硝态氮的去除率最高分别可达97.86%、91.32%和87.72%。综合分析菌株L12的氮源利用性能、中间产物及气态产物、氮平衡、脱氮酶活性和功能基因鉴定结果推测菌株L12在含La(Ⅲ)条件下的同步硝化反硝化(SND)脱氮途径为NH4+-N→NH2OH→NO2--N→NO3--N→NO2--N→NO→N2O→N2。菌株L12在含镧废水中具有优良的脱氮特性,表明了其在稀土尾矿废水脱氮中的应用潜力。
稀土是化学周期表中镧系元素和钪、钇等共17种金属元素的总称,包含轻稀土和重稀土两类。从下游来看,稀土产品应用于强磁材料、箱包玩具、陶瓷、音响、汽车、催化、照明等领域,用途广泛。目前,离子型稀土矿的提取主要采用原地浸矿工艺,该工艺是利用浸矿剂(主要是硫酸铵)中的氨氮与稀土矿中的稀土离子发生离子交换反应,使稀土组分进入浸出液,再进行工艺处理最后得到稀土氧化物。然而,在这个过程中,大量浸矿剂残留在矿山和土壤中,通过雨水等流入水体中,产生大量难以处理的稀土矿山氨氮废水。氮素超标时,会引起水体富营养化,破坏水体生态平衡,严重威胁动植物生长及人类健康。
生物脱氮法是目前应用最广泛的氮污染控制方法。然而,稀土矿山废水中除含有大量的含氮物质外,还含有大量的稀土元素离子。据报道,靠近稀土矿井的溪流中稀土元素的总含量甚至超过20mg/L。高浓度稀土离子对菌株会产生严重抑制效果。董炎等处理高氨氮稀土废水,在La(Ⅲ)浓度小于1mg/L时氨氮去除率为85.99%,La(Ⅲ)浓度大于1mg/L时厌氧氨氧化菌活性受到了严重抑制,氨氮去除率不足70.00%。李芸等研究发现0~10mg/L的Er(Ⅲ)对硝化细菌(AOB)活性呈现促进作用,浓度20mg/L以上的Er(Ⅲ)对AOB活性呈现抑制作用,在60mg/L时AOB活性降低了31.79%。Xiao等使用宏基因组测序技术检测了不同Ce(Ⅲ)浓度作用下氮代谢微生物和相关基因的丰度,发现AOB和反硝化细菌(DNB)活性受高浓度Ce(Ⅲ)抑制。上述研究进一步表明稀土元素离子会显著影响微生物生长及脱氮过程。
同步硝化反硝化(SND)技术可以在单一有氧条件下同时进行硝化和反硝化反应,不仅节省了反应器空间和能源,也降低了与pH调整相关的成本,越来越受到人们的关注。异养硝化-好氧反硝化(HN-AD)菌是SND技术的关键功能菌种,如微小杆菌(Exiguobacterium)H1和不动杆菌属(Acinetbacter sp.)的ZH7等,而且有研究发现HN-AD菌可以在低温、高温、高盐、高酸碱性和高重金属浓度等多种极端环境中生长和保持较高的脱氮效率。当Cu(Ⅱ)/Zn(Ⅱ)浓度高达80mg/L时,恶臭假单胞菌(Pseudomonas putida)ZN1仍能去除97.79%的氨氮,贪铜菌属(Cupriavidus sp.)的S1可以在Zn(Ⅱ)浓度为20mg/L条件下去除97.42%的氨氮。稀土矿山废水含有大量的稀土元素,但目前关于稀土元素对HN-AD菌脱氮性能影响的研究总体上还比较匮乏。因此,有必要开展稀土金属元素对HN-AD菌生长及脱氮性能的影响,以便为SND技术用于处理稀土尾矿废水提供优良菌种资源。
本实验从实验室地下水中分离筛选出一株能在含有La(Ⅲ)环境中生长,并具有优异氨氮去除性能的菌株。首先,采用单因素实验探究了碳源、C/N、温度、pH、氨氮浓度和La(Ⅲ)浓度等不同环境因素对菌株在含La(Ⅲ)环境中硝化性能的影响。然后,通过不同氮源利用情况研究了菌株在含La(Ⅲ)环境中的异养硝化和好氧反硝化性能。最后,结合中间产物、氮平衡、关键酶活性以及基因组框架图测序分析结果,研究菌株在镧离子胁迫下的脱氮路径和机理,不仅可为稀土污水脱氮处理提供重要菌种资源,还对丰富和发展特殊条件下的生物脱氮理论有重要的科学意义。
LB培养基(1L):胰蛋白胨5g,酵母浸粉5g、NaCl 10g,调节pH为7.0。
选择(SM)培养基(1L):CH3COONa 4.783g、(NH4)2SO4 0.472g、K2HPO4 0.25g、NaCl 0.25g、1mL微量元素溶液,调节pH为7.0。
硝化(NM)培养基(1L):碳源C4H4Na2O4 4.05g、La(Ⅲ) 10mg/L,其余成分同SM培养基,调节pH为7.0。
反硝化(DM)培养基(1L):氮源NaNO3 0.61g(或者NaNO2 0.49g)、La(Ⅲ) 10mg/L,其余成分同NM培养基,调节pH为7.0。
微量元素溶液(1L):MgSO4·7H2O 0.5g、MnSO4·H2O 0.1g、FeSO4·7H2O 0.2g。培养基中La(Ⅲ)由对应体积的LaCl3·7H2O溶液[2.67g LaCl3·7H2O溶于100mL高纯水,La(Ⅲ)浓度10g/L]提供。在液体培养基基础上加入0.018kg/L琼脂制备固体培养基。所有培养基均需要用高压蒸汽灭菌锅在122℃灭菌20min。
将实验室地下水5mL加入100mL无菌LB培养基中,在30℃、120r/min条件下培养48h以获得富集菌悬液。取1mL上述菌悬液转移到含10mg/L La(Ⅲ)的SM培养基中培养48h,继续将菌液依次转移至含50mg/L和100mg/L La(Ⅲ)的SM培养基中相同条件继续驯化(每种浓度重复3次)。取最后一次驯化后菌液梯度稀释10-4倍在固体SM培养基平板上涂布,30℃生物培养箱中培养72h。
在超净工作台中用无菌接种环挑出形态不同的单菌菌落在无菌固体SM培养基平板上划线纯化,将纯化后的单菌菌落接种到含100mg/L La(Ⅲ)的无菌SM培养基中,在30℃、120r/min条件下培养60h后测试其生长及氨氮去除性能,选择脱氮性能最强的菌株为目标菌株用于后续实验研究。最后,将目标菌株纯化3次后,接种于NM培养基在30℃、120r/min条件下培养至对数期,于超净工作台中取对数期菌悬液0.5mL与甘油1∶1(体积比)混合保存于-80℃超低温冰箱中。
将筛选的菌株接种到NM固体培养基中,生物培养箱30℃培养36h,观察菌株形态。将培养的菌株送至艾基生物技术公司提取细菌基因组DNA,采用通用引物27F(5′-AGAGTTTGATCMTGGCTCAG-3′)和1492R(5′-TTGGYTACCTTGTTACGACT-3′)扩增16S rDNA序列并测序。通过BLAST将测得的基因序列与美国国家生物技术信息中心(NCBI)Genbank中其他微生物的序列进行对比,然后通过MEGA 11软件采用邻接法(neighbor-joining)来构建菌株的系统发育树。
采用单因素控制变量法分别从不同碳源(乙酸钠、葡萄糖、柠檬酸钠、丙酮酸钠、丁二酸钠)、C/N比(8、10、12、14、16、18)、初始pH(5、6、7、8、9、10、11)、温度(15℃、20℃、25℃、30℃、35℃、40℃)、初始氨氮浓度(0、50mg/L、100mg/L、200mg/L、500mg/L、1000mg/L)、La(Ⅲ)浓度(0、5mg/L、10mg/L、25mg/L、50mg/L、75mg/L、100mg/L、200mg/L)这6个方面研究不同环境因素对菌株L12硝化性能的影响。每组实验设置3组平行样,将培养9h处于对数期的菌株L12的菌悬液(1mL)接种到La(Ⅲ)浓度为10mg/L的100mL NM培养基中,培养60h。除改变单一变量元素外,其他培养条件均为30℃、120r/min、10mg/L La(Ⅲ)和初始pH为7.0。每组实验进行期间,每隔12h取菌悬液,测定其OD600(溶液在600nm波长处的吸光值),离心(10000r/min,5min)后取上清液,测定其氨氮(NH4+-N)、亚硝态氮(NO2--N)、硝态氮(NO3--N)、总氮(TN),化学需氧量(COD)的浓度和pH。
取培养9h处于对数期的菌株L12的菌悬液1mL,分别接种于以氨氮为唯一氮源的NM培养基(100mL)和以硝酸钠或亚硝酸钠为唯一氮源的DM培养基(100mL)中,在30℃、初始pH为8.0和120r/min条件下培养60h,每隔12h取菌悬液,测定其OD600,离心(10000r/min,5min)后取上清液,测定其氨氮、亚硝态氮、硝态氮、总氮,COD的浓度和pH。
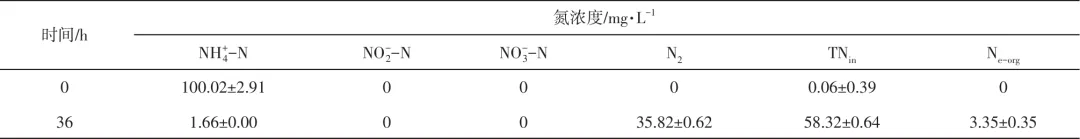
在含100mL NM培养基[La(Ⅲ)浓度为10mg/L]的250mL气密瓶中,接种1%活化后的菌液后充满高纯度氧气,然后密封瓶口,30℃、120r/min培养36h,收集0h和36h瓶内气体用于N2分析,然后收集菌液,测量其总氮TN1的浓度,离心(10000r/min,5min)后取上清液,测量其氨氮、亚硝态氮、硝态氮和总氮TN2的浓度。根据式(1)、式(2)计算细胞内氮(TNin)和细胞外有机氮(Ne-org)的浓度。
以1%接种量将处于对数期的菌株L12菌悬液接种于La(Ⅲ)浓度为10mg/L的NM培养基中,在30℃、120r/min条件下培养,18h时取菌液离心(10000r/min,10min),用磷酸盐缓冲盐溶液(PBS)清洗、重悬,重复3次后冰浴条件下超声波粉碎20min(工作7s,间隔4s),收集粗酶液。蛋白质含量用考马斯亮蓝法测定,氨单加氧酶(AMO)活性采用氨单加氧酶检测试剂盒(江苏酶免实业公司)测定,羟胺氧化还原酶(HAO)、亚硝酸盐还原酶(NIR)和硝酸盐还原酶(NAR)活性通过反应中羟胺、亚硝酸盐和硝酸盐的减少量测定。
将培养菌株L12送至伟寰生物科技有限公司(上海),提取细菌基因组DNA后,基于Illumina HiSeq平台进行基因组框架测序。测序的结果经过过滤、组装和优化,结合NR、GO和KEGG数据库中与N代谢相关的功能基因及代谢通路对菌株L12进行基因预测和功能注释。
采用紫外-可见分光光度计在波长600nm处测得的吸光度来反映菌株的生长情况。水中氨氮、亚硝态氮、硝态氮和总氮、COD的浓度分别用纳氏试剂分光光度法(HJ 535—2009)、N-(1-萘基)-乙二胺分光光度法(GB 7493—1987)、水杨酸-浓硫酸分光光度法(HJ/T 346—2007)、碱性过磺酸钾消解紫外分光光度法(HJ 636—2012)和重铬酸钾-硫酸银法(HJ 828—2017)测定。采用pH计(FE28,梅特勒)测定pH。氨氮、硝态氮、亚硝态氮以及COD去除率η的计算方法如式(3)所示。
式中,c0为初始氨氮浓度、硝态氮浓度、亚硝态氮浓度以及COD值,mg/L;ct为培养至t时间时对应的氨氮浓度、硝态氮浓度、亚硝态氮浓度及COD值,mg/L。
所有实验重复3次,实验结果主要以平均值±标准差的形成表示,数据均使用Origin 2021软件进行单因素方差分析和作图,以p<0.05作为显著性差异的标准。
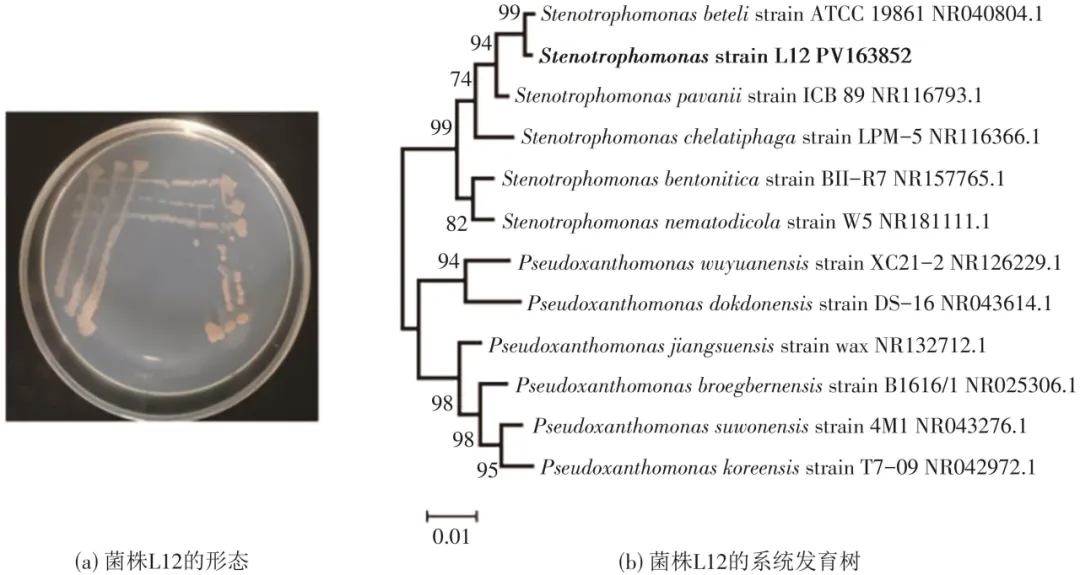
从废水中经富集、驯化和分离后,得到一株耐镧异养硝化-好氧反硝化细菌,命名为L12。从图1(a)中可知,菌株L12在固体NM培养基上显示为浅红色,菌落呈圆形,表面平滑,边缘规则。采用16S rDNA通用引物成功扩增16S rDNA,经测序获得长度约为1540碱基对的目标基因片段。将该片段提交至GenBank数据库,得到菌株L12的基因序列号为PV 163852。利用基于局部比对搜索工具(BLAST)对菌株L12的16S rDNA序列进行同源性序列比对,构建的系统发育树如图1(b)所示。发现其与寡养单胞菌属(Stenotrophomonas sp.)多个成员关系密切,其中与Stenotrophomonas beteli ATCC19861(登录号:NR 040804.1)相似度最高,达99.57%,故初步判定该菌株为寡养单胞菌属。该菌属已被报道对金属如Cd、Zn有一定耐受作用,因此有必要开展对菌株S. L12在含镧离子废水中的脱氮性能的相关研究。
图1 菌株L12的形态和16S rDNA系统发育树
La(Ⅲ)存在下不同环境因素对菌株L12氨氮降解性能的影响
2.2.1 碳源
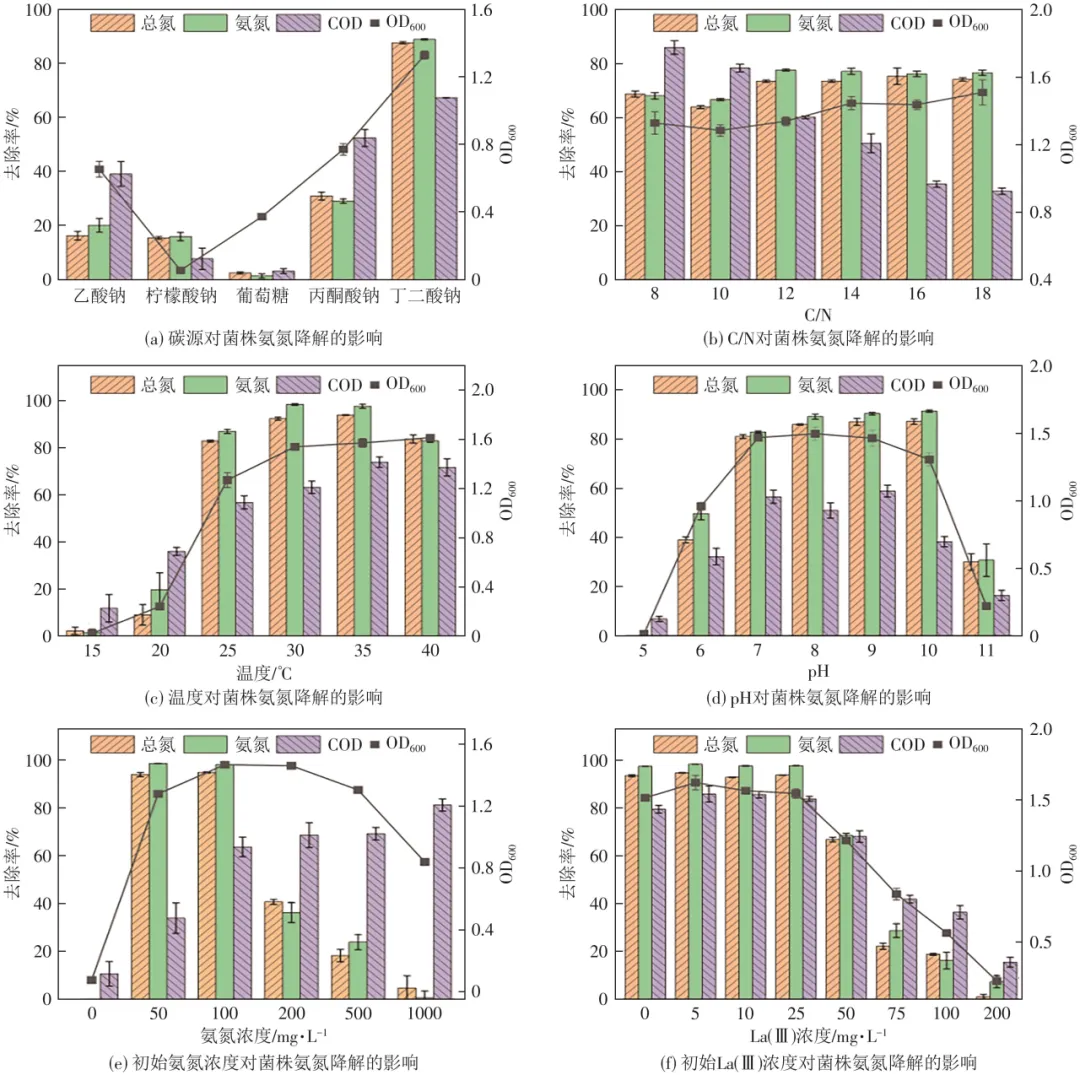
如图2(a)所示,菌株L12能够以乙酸钠、丁二酸钠和丙酮酸钠为唯一碳源进行生长。这些碳源比较容易被细菌利用,因为它们参与了三羧酸(TCA)循环,产生更多的还原态烟酰胺腺嘌呤二核苷酸(NADH)和腺嘌呤核苷三磷酸(ATP),电子传递系统活性(ETSA)较高。而当以丁二酸钠为唯一碳源时,菌株L12的OD600、氨氮和总氮去除率均达到最高(p<0.05),分别为1.33、88.92%和87.67%。因此,在研究范围内选择丁二酸钠作为菌株L12的最佳碳源,这与许多异养硝化-好氧反硝化菌选择相一致。
图2 不同环境因素对菌株L12在含La(Ⅲ)环境中氨氮去除性能的影响
2.2.2 C/N比
C/N比对菌株L12生长及氨氮利用性能的影响如图2(b)所示,当C/N比不高于12时,菌株L12的氨氮去除性能随着C/N比增加呈现先略微下降后显著升高的变化趋势,C/N比为12时氨氮去除率最高(p<0.05),为77.74%。之后继续升高C/N比,氨氮去除率基本保持不变(p>0.05),这是因为细菌的HN-AD性能是有限的,达到最高水平后继续提高C/N比并不会继续提高硝化过程中微生物的代谢和电子传递速率。
2.2.3 温度
如图2(c)所示,低温下(15~20℃)菌株生长和氨氮去除性能受到明显抑制,20℃时菌株氨氮去除率仅为19.74%。温度升高至25℃时,菌株延滞期缩短,24h时OD600达到1.27,氨氮去除率为86.94%。30℃和35℃菌株生长最为快速,24h的OD600达到1.54和1.57,同时氨氮和总氮去除率显著高于其他温度(p<0.05),分别为98.40%和97.69%。而40℃时温度过高,脱氮酶活性也会受到抑制,24h时的氨氮去除率有所降低,为82.91%。综上,30~35℃为菌株的最佳生长温度范围,这与之前报道的大多数HN-AD细菌最适温度范围接近。
2.2.4 初始pH
如图2(d)所示,初始pH为5和11时,菌株基本不生长,氨氮浓度基本不变,这说明pH过高或者过低都会抑制菌株生长及脱氮性能。初始pH为8~10时,菌株OD600均不低于1.31,硝化能力最强,氨氮去除率均在90%以上。总体来说,弱碱性条件下的菌株L12生长速度最快,且氨氮去除率略高于中性(pH 7,82.94%),远高于酸性(pH 6,49.79%)。在其他HN-AD细菌中也观察到这一趋势,可能是因为碱性环境中存在游离氨,从而促进了异养硝化过程。
2.2.5 初始氨氮浓度
图2(e)为初始氨氮浓度对菌株L12生长及氨氮去除的影响。缺少氮源(初始氨氮浓度为0)时,菌株L12基本不生长。当初始氨氮浓度为50mg/L和100mg/L时,菌株L12生长最快,氨氮处理效果最佳,24h氨氮去除率分别达到98.59%和98.02%。当初始氨氮浓度升高至200mg/L和500mg/L,菌株L12虽然仍保持生长,但生长速度减缓,氨氮去除率更是降至36.28%和23.97%,且24h之后氨氮也基本没有继续去除(图中数据未显示)。当初始氨氮浓度继续升高至1000mg/L,24h时的OD600仅有0.84,且氨氮基本没有去除,这可能是由碳源不足和高浓度氨氮的毒性所致的。综合考虑菌株L12生长及氨氮去除性能,将初始氨氮浓度固定为100mg/L进行后续实验。
2.2.6 不同La(Ⅲ)浓度
从图2(f)中可以看出,与不添加La(Ⅲ)的对照组相比,添加5mg/L La(Ⅲ)对菌株L12生长表现出良好的促进作用。24h时OD600和氨氮去除率(1.62和98.30%)均高于没有La(Ⅲ)添加时(1.52和97.44%),与之前研究结果一致。当La(Ⅲ)浓度达到10mg/L和25mg/L时,细菌开始通过增强膜转运,将过量的镧转运到细胞外,以及产生更多的胞外聚合物(EPS),以降低镧的毒性,此时菌株生长和氨氮去除效果未受到La(Ⅲ)毒性显著影响,与不添加La(Ⅲ)时接近。但当La(Ⅲ)浓度达到50mg/L、75mg/L和100mg/L时,膜转运和EPS的效果会被抑制,进而抑制细菌生长(OD600分别为1.22、0.84、0.57),氨氮去除率相比10mg/L时分别下降了29.84%、70.55%和83.36%。当La(Ⅲ)浓度达到200mg/L,菌株生长基本停滞,此时对应氨氮去除率仅为7.34%。综上所述,0~25mg/L的La(Ⅲ)对菌株L12生长和脱氮影响不大;La(Ⅲ)浓度大于25mg/L,会对菌株的生长和脱氮造成明显影响,且La(Ⅲ)浓度越高对菌株生长和氨氮去除能力抑制越显著。
重金属离子可以诱导微生物DNA单链断裂、碱基错配,干扰功能基因表达,对微生物去除氨氮有重大影响,如表1所示。如Zn(Ⅱ)暴露减少了酰基高丝氨酸内酯(AHLs)的分泌,影响了EPS和氧化还原物质的分泌,降低了细胞外电子传递效率;同时还导致与脱氮相关的酶活性下降,改变细胞膜通透性,导致微生物能量代谢受阻。研究表明,在8mg/L和20mg/L Zn(Ⅱ)下,气单细胞菌属(Aeromonas sp.)HN-02和Cupriavidus sp. S1的氨氮去除率只有94.59%和97.42%。两株耐重金属菌株在100mg/L Zn(Ⅱ)下,氨氮去除率低至88.75%和84.60%,且去除时间长达48h。Cd(Ⅱ)毒性较大,假单胞菌属(Pseudomonas sp.)XF-4需要36h才能在10mg/L Cd(Ⅱ)环境下去除30mg/L氨氮。与其他重金属类似,稀土金属对细胞也存在毒性。胡锦刚等从江西稀土矿周边土壤中分离出1株HN-AD菌黄褐假单胞菌(Pseudomonas fulva)K3,在初始氨氮浓度为100mg/L条件下平均氨氮去除速率为4.03mg/(L·h)。本菌株在浓度0~25mg/L的La(Ⅲ)下仍保持正常生长,异养硝化基本不受影响。
表1 不同菌株在含不同重金属离子环境中的氨氮去除性能比较
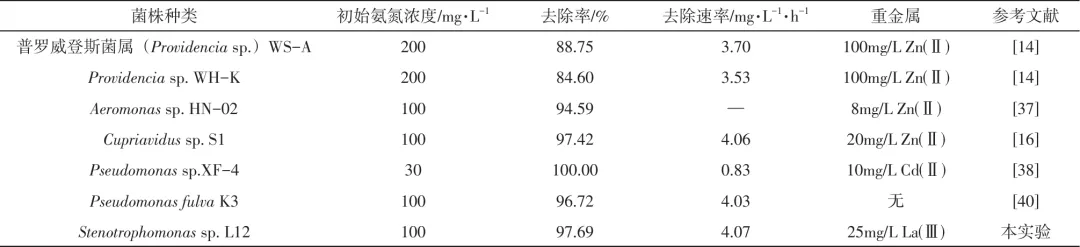
在前文中探究了不同环境因素对菌株L12在含La(Ⅲ)环境下的氨氮降解性能的影响,得出菌株L12在10mg/L La(Ⅲ)环境下最优氨氮降解条件为碳源为丁二酸钠、C/N为12、培养温度为30℃、初始pH为8。后续本文将在上述最优条件下系统研究菌株L12在含La(Ⅲ)环境下的脱氮性能。菌株L12以氨氮为唯一氮源的脱氮过程如图3所示。前6h菌株处于适应期,生长比较缓慢,氨氮浓度缓慢降低至95.77mg/L,适应期比优化之前明显缩短。6~24h菌株处于对数期,OD600值从0.03迅速增长到1.53,氨氮主要在这个阶段被迅速降解,24h已基本去除,去除率达到97.86%,高于HN-AD菌如Pseudomonas putida DS2的(91%)和卓贝尔氏菌(Zobellella)B307(96%)的氨氮去除率,其硝化过程中未出现明显的硝态氮和亚硝态氮积累。总氮和COD去除趋势与氨氮去除类似,最大去除率均发生在对数期,24h时总氮去除率为91.88%,COD去除率为83.85%。24~48h菌株进入稳定期,OD600稳定在1.6左右,氨氮浓度稳定在1.5mg/L左右。培养48h后,菌株进入衰亡期,氨氮浓度上升至2.82mg/L,OD600也略有下降,COD值已不足200mg/L,这可能是碳源不足引起菌株衰亡所致的。硝化过程中pH稳步上升,从初始的8.0上升到60h的9.2,过程中产生的弱碱性环境更适合菌株生长。
图3 La(Ⅲ)存在下菌株L12以氨氮为唯一氮源的异养硝化过程
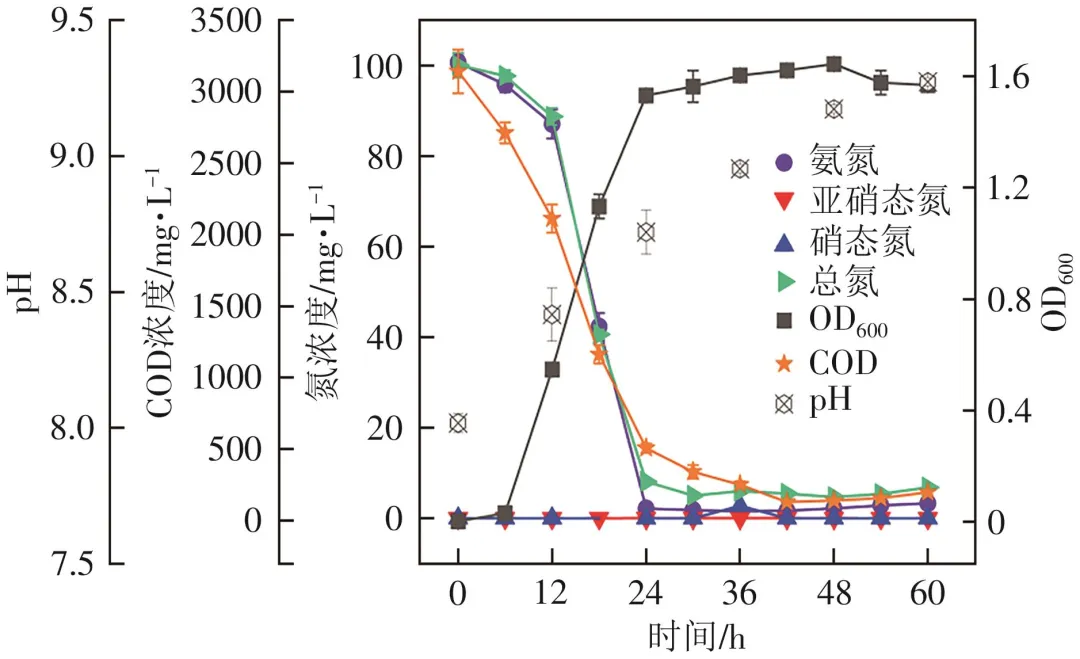
好氧反硝化主要指微生物在好氧条件下以有机物为碳源和电子供体,以氧气和硝态氮为电子受体,将硝态氮还原为氮气的酶促反应过程。首先,考察了La(Ⅲ)存在下菌株L12以硝酸钠为唯一氮源时的好氧反硝化性能。如图4所示,与氨氮降解过程类似,前6h菌株L12处于延滞期,生长缓慢。培养6~24h时处于对数期,菌株L12的OD600从0.01迅速增加至1.46,与此同时硝态氮浓度从6h的98.11mg/L降低到24h的38.09mg/L,12~24h的硝态氮平均去除速率最高,为4.75mg/(L·h)。18h时积累了13.33mg/L亚硝态氮,之后逐渐降低。推测菌株L12能利用亚硝态氮进行好氧反硝化。氨氮浓度无明显变化,一直维持在接近为零的状态。培养24h后菌株L12进入稳定期,48h时OD600达到峰值1.55,硝态氮、总氮和COD均达到最高去除率91.32%、88.47%和95.94%,之前积累的亚硝态氮也被基本消除。48h以后菌株进入衰亡期,OD600开始下降,硝态氮浓度出现反弹。
图4 La(Ⅲ)存在下以硝酸钠为唯一氮源时菌株L12的生长和反硝化性能
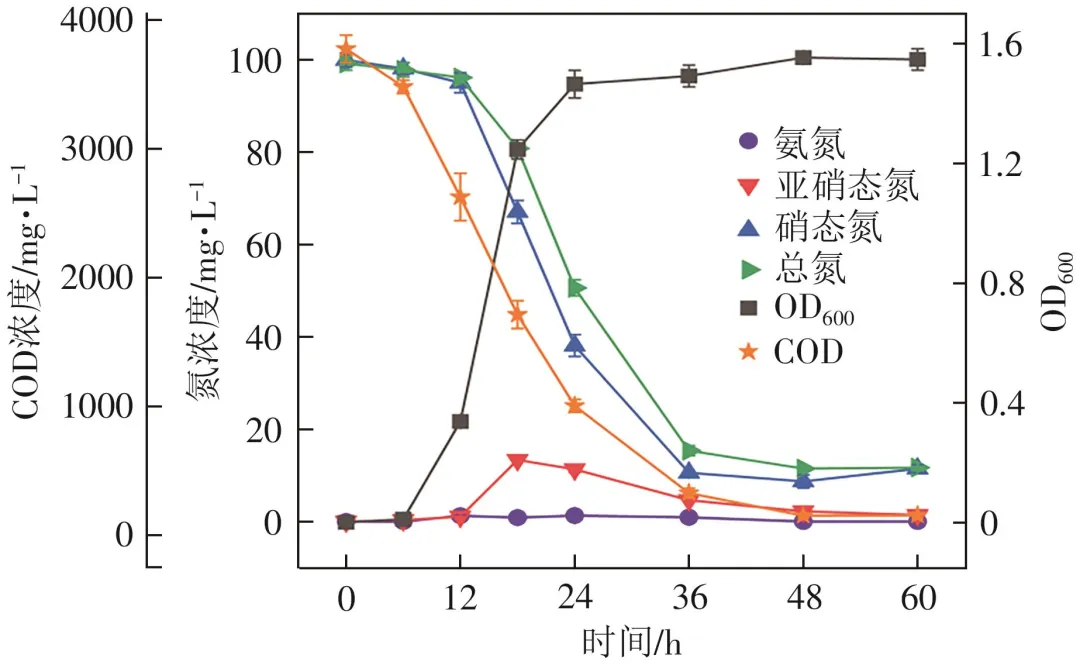
在La(Ⅲ)存在下菌株L12以亚硝酸钠为唯一氮源时的好氧反硝化性能见图5。菌株培养前6h OD600基本不变,此阶段为菌株的生长停滞期;6~24h菌株进入快速生长期,OD600从0.02增加至1.50,亚硝态氮浓度从97.23mg/L降低到40.55mg/L,18h时积累了7.73mg/L的硝态氮,之后被逐渐去除。24h以后菌株进入稳定期,36h时OD600达到峰值1.55,亚硝态氮、总氮和COD去除率可达85.55%、84.43%和92.67%。48h时亚硝态氮去除率最高,为87.72%。此外,亚硝态氮为唯一氮源时产生氨氮要多于以硝态氮为唯一氮源产生的,这可能与亚硝态氮的毒性导致细菌死亡有关。综上可得,菌株L12在La(Ⅲ)存在下能够以硝态氮和亚硝态氮为唯一氮源进行生长和好氧反硝化,反硝化效果不如硝化可能是因为菌株相对更容易利用氨氮作氮源,这一现象在其他HN-AD菌株中也有出现。
图5 La(Ⅲ)存在下以亚硝酸钠为唯一氮源时菌株L12的生长和反硝化性能
上述研究结果表明,菌株L12在含La(Ⅲ)条件下能够分别以氨氮、硝态氮和亚硝态氮为唯一氮源进行生长,推测菌株L12具有异养硝化-好氧反硝化性能。随后,采用气相色谱检测菌株L12脱氮过程中的气态产物,结果发现菌株L12在含La(Ⅲ)条件下以氨氮为唯一氮源时可以生成氮气,进一步证明菌株L12的好氧反硝化性能。表2为氮平衡实验结果,表明菌株L12以氨氮为唯一氮源生长时有98.34%的氨氮被利用,其中约58.32%氨氮被同化为细胞内氮,35.82%转化生成氮气,3.35%转化为胞外的有机氮,基本没有硝态氮和亚硝态氮的积累,表明菌株L12在含La(Ⅲ)条件下氨氮代谢过程主要包括同化为有机氮,以及经异养硝化-好氧反硝化过程转变成氮气两种路径,这与大部分异养硝化-好氧反硝化菌脱氮过程类似。
表2 La(Ⅲ)存在下菌株L12在以氨氮为唯一氮源异养硝化过程中的氮平衡
在SND过程中,通常测定几种关键酶AMO、HAO、NAR和NIR的活性用来分析细菌的氮代谢途径。其中,AMO负责将氨氧化为羟胺,HAO将羟胺氧化为亚硝酸盐,而NAR和NIR分别负责硝态氮和亚硝态氮的反硝化过程。如表3所示,菌株L12在含La(Ⅲ)条件下的氨氮利用过程中,4种脱氮关键酶活性均被检测出来,HAO、AMO酶比活力为0.082U/mg protein和0.134U/mg protein(U为每分钟催化1μmol底物反应的酶量,mg protein为每毫克蛋白中的酶活性),高于斯氏假单胞菌(Pseudomonas stutzeri)ADP-19的HAO(0.020U/mg protein)和AMO(0.011U/mg protein)的酶比活力;NAR、NIR酶比活力为0.045U/mg protein和0.031U/mg protein,远高于根瘤菌属(Rhizobium sp.)WS7的NAR(0.006U/mg protein)和NIR(0.002U/mg protein)的酶比活力,从酶活水平证明菌株L12具有异养硝化-好氧反硝化性能。
表3 菌株L12在含La(Ⅲ)条件下的氨氮利用过程中脱氮关键酶活性
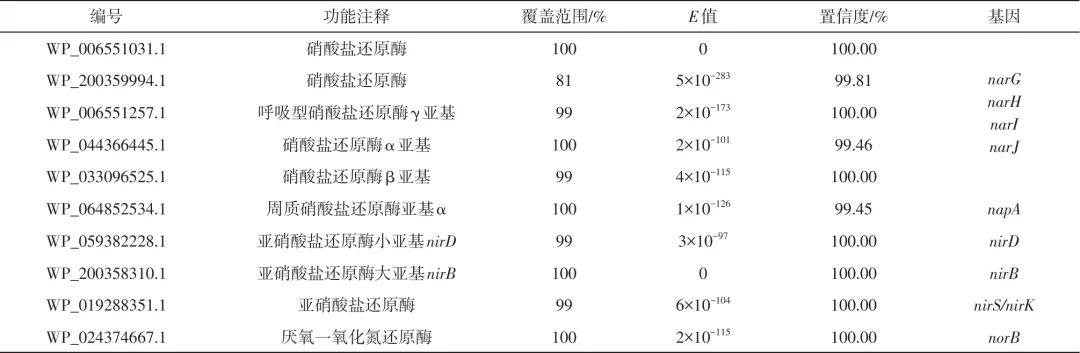
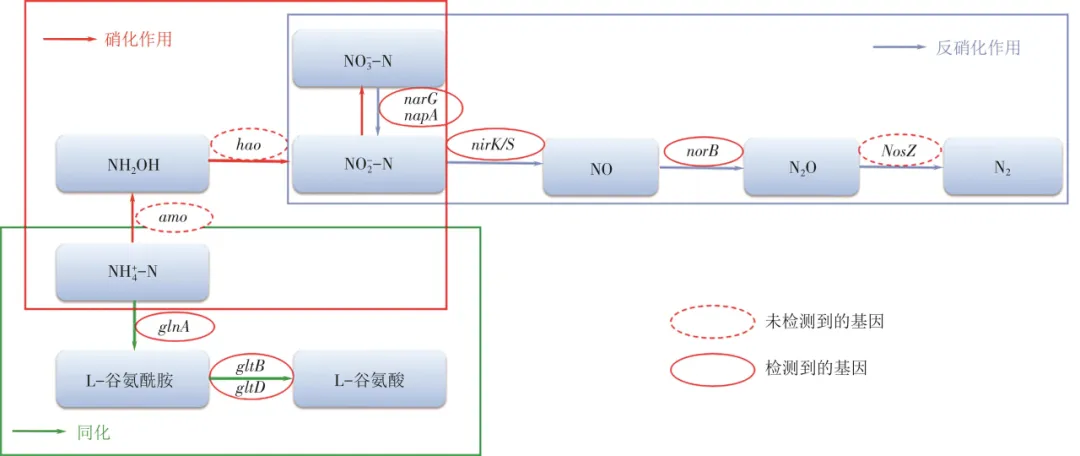
为深入研究菌株L12在含La(Ⅲ)条件下的脱氮路径,对其基因组框架进行测序,将获得的结果在美国国家生物技术信息中心(NCBI)提供的BLASTx与NR数据库中用脱氮相关蛋白编码基因进行比对和注释,其结果如表4所示。在菌株L12基因组序列中检测到多个参与反硝化过程的关键酶基因,其中包括编码呼吸型硝酸盐还原酶(Nar)的功能基因narG/narH/narI/narJ、编码周质硝酸盐还原酶(Nap)的功能基因napA、编码异化型亚硝酸盐还原酶的功能基因nirS/nirK、同化型亚硝酸盐还原酶的功能基因nirB/nirD和编码一氧化氮还原酶(Nor)的功能基因norB。这些反硝化关键酶基因的存在,为菌株L12在含La(Ⅲ)条件下的异养硝化-好氧反硝化脱氮路径提供了基因水平上的证据。菌株基因组框架序列中并未检测到硝化相关的amo和hao基因,这在克雷伯氏菌属(Klebsiella sp.)KSND和克雷伯氏肺炎杆菌(Klebsiella pneumoniae)EGD-HPI9-C等HN-AD菌株中也发现了类似现象,可能是因为存在目前未知的酶与AMO/HAO酶功能相同,其可以氧化氨和羟胺,传统硝化的路径有待进一步完善。
表4 菌株L12基因组中与反硝化关键酶相关的蛋白编码基因
综合分析菌株L12在含La(Ⅲ)条件下的氮源利用性能、产气及氮平衡以及脱氮关键酶活性和其相关编码基因,结果表明菌株L12主要通过同化为胞内氮和异养硝化-好氧反硝化产生氮气两条路径利用氨氮,其中以氨氮为唯一氮源的异养硝化-好氧反硝化的SND脱氮途径可能为NH4+-N→NH2OH→NO2--N→NO3--N→NO2--N→NO→N2O→N2(图6),这与许多异养硝化-好氧反硝化细菌脱氮路径相一致。此外,根据KEGG代谢通路注释,菌株L12的氨氮同化途径中还检测到编码谷氨酰胺合成酶的基因glnA和谷氨酸合成酶的基因gltB/gltD。谷氨酰胺合成酶可以将NH4+-N转化为L-谷氨酰胺,谷氨酸合成酶可以将L-谷氨酰胺转化为L-谷氨酸。因此,推测菌株L12以氨氮为唯一氮源的同化路径可能为NH4+-N→L-谷氨酰胺→L-谷氨酸(图6)。
(1)筛选出一株在La(Ⅲ)胁迫下高效SND的菌株Stenotrophomonas sp. L12。当初始La(Ⅲ)浓度为10mg/L和氨氮浓度为100mg/L时,菌株L12利用氨氮的最佳条件为碳源为丁二酸钠、C/N比为12、30℃和pH为8.0。此外,菌株L12在含La(Ⅲ)环境中还能够在50~500mg/L氨氮浓度范围内生长和利用氨氮。
(2)菌株L12表现出较强的La(Ⅲ)耐受性。在La(Ⅲ)浓度为0~25mg/L条件下24h时氨氮去除率达到97%以上,COD去除率达到80%以上。但当La(Ⅲ)浓度大于25mg/L,菌株L12生长和脱氮效率受到明显抑制。
(3)氮平衡表明大部分氨氮被同化,剩余的氨氮基本通过SND作用转化为气态氮。结合菌株L12氮源利用性能、氮平衡、脱氮酶活性和基因组功能注释等结果推测菌株L12的SND脱氮路径为NH4+-N→NH2OH→NO2--N→NO3--N→NO2--N→NO→N2O→N2。
第一作者:陈虎,博士,讲师,研究方向为废水生物脱氮。
通信作者:王莹,讲师,研究方向为废水污染物生物处理。
邮发代号:82-311
订阅热线:010-64519502
网址:http://www.hgjz.com.cn